Look at the trees! Evolution and engineering in action
A short essay on trees from an engineering perspective, and its implications for evolution and design
This post comes from a bit of wikipedia rabbit-hole-diving I did while brushing up on the theory of buckling in columnar structures for work. It started with some curious fun facts about the way trees grow and ended up revealing many interconnected threads that demonstrate the power of evolution. Consilience (convergence of independent lines of evidence on a singular conclusion) tends to do that, of course.
Why do trees grow tall?
Thermodynamics of trees
Every living thing requires energy to survive, grow and reproduce. The Gibbs free energy $ G $ quantifies the amount of free (available) energy from any closed system at constant temperature and pressure (a good model of most chemical processes). For any spontaneous chemical reaction, the change in this free energy, $ \Delta G $, must be negative, or else external energy must be supplied to facilitate the process, and if this energy source is removed, the process will become spontaneous in the opposite direction.
Plants have embarked on the thermodynamically-Sisyphean task of getting carbon dioxide to do stuff for them, rolling carbon-containing molecules up the Gibbs free energy mountain (the Calvin cycle of photosynthesis) to make food molecules (glucose and carbohydrates) which release the energy back to them (respiration), before starting over. In human-centric life, we think of CO$_2$ as a waste product that’s good for nothing except anthropogenic global warming: it’s the end product of both our individual respiration and our species’ industrial-scale combustion of fossil fuels, not a starting point.
. Data sourced from Figure 6 of ([Jiang *et al.*, 2010](https://www.researchgate.net/publication/44691024_Turning_carbon_dioxide_into_fuel)) and [Wikipedia](https://en.wikipedia.org/wiki/Standard_Gibbs_free_energy_of_formation).](/post/look-at-the-trees/gibbs-free-energy-mountain_hub438ae23bf39ed0aead7d219643bfc73_559515_565e425173dc5b0aa8b82d05620cb960.webp)
Plants aren’t fighting entropy alone: they get away with using CO$_2$ as their input because they have the Sun, beaming in limitless free energy to a planet that would otherwise be isolated and lifeless. Of course, it wasn’t plants who came up with this trick. That goes back to the original photosynthetic microorganisms, a phylum of prokaryotes called cyanobacteria, which found a way to make chemical fuel (ATP and NAPDH) from visible photons for use in its own metabolism. The ATP stores the free energy because ATP hydrolysis and NAPDH oxidation are both exergonic and can be coupled to facilitate endergonic enzymatic reactions:
$$ ATP + H_2O \xrightarrow[]{\Delta_f G’ ^{\ominus} \ = \ -37 \ kJ/mol} ADP + P_i $$
$$ NAD(P)H + H^+ + \frac{1}{2} O_2 \xrightarrow[]{\Delta_f G’ ^{\ominus} \ = \ -220 \ kJ/mol} NAD(P)^+ + H_2O $$
After throwing dozens of ATPs and NADPHs and half a dozen carbon dioxide and water molecules into various enzyme complexes, the free energy balance shifts just enough to pull us up the mountain and make a single molecule of precious glucose:
$$ 6 \ CO_2 \ + \ 6 \ H_2O \xrightarrow[]{\Delta_f G’ ^{\ominus} \ = \ +2,872 \ kJ/mol} C_6H_{12}O_6 \ + \ 6 \ O_2 $$
with the thermodynamic debt having been paid for in the currency of ATP. Ancient protists, like the ancestors of algae, just took control of this mini-generator and put it into an organelle (endosymbiosis) that we now call the chloroplast.
, which is then fed into the (light-independent) Calvin cycle above to fix carbon dioxide into larger organic molecules like GA3P (glyceraldehyde 3-phosphate) and later glucose and polysaccharides like starch and cellulose. Since the RuBisCo enzyme will happily bind oxygen nearly as easily as carbon dioxide, wasting energy, three variants of carbon capture have evolved in plants (C3, C4 and CAM) as a defence against [photorespiration](https://en.wikipedia.org/wiki/Photorespiration) in their specific climates.](/post/look-at-the-trees/calvin-cycle_hu5a5ca89b72602000bb74a574b18a5973_623124_378df6d4cb7d458b96a751936e0a1bba.webp)
Ecology of trees
Once multicellular, plant evolution was guided by one rule:
$$ \text{more sunlight → more energy → more growth → more sunlight.} $$
It doesn’t matter that after several billion years of evolution, the round-trip energy efficiency of photosynthesis is barely more than 1%, a figure that pales in comparison to manmade semiconductor-based solar photovoltaic cells that are now pushing 30% efficiency after merely a few decades of technological progress. Any utilisation of an omnipresent energy influx like sunlight is better than none. For trees, since more growth also comes with more reproduction via further seed dispersal and germination, this positive feedback became subject to natural selection.
The problem is, trees don’t grow in a vacuum. When seeds fall to land, they tend to produce multiple plants in close proximity - you never see a lone blade of grass, for example. That means a battle for the soil’s finite sources, as well as reduction in an individual plant’s sunlight exposure due to shadows cast by the others. This is the recipe for Malthusian competition, which underpins the reason behind classical Darwinian natural selection (the ‘struggle for existence’). Unless you opt for a symbiotic strategy, growing taller than your conspecifics becomes essential to survival, and this directional selection is illustrated in the plant fossil record.
How do trees grow tall?
Biomaterials like wood are not known for their structural homogeneity, and therefore develop imperfections easily. Structurally, this means lower safety factors, and earlier onset of structural instability (for buckling, this is formalised by Perry’s analysis*, later incorporated into standard engineering design codes based on experiments by Robertson in the 1920s.). Trees therefore must respond to structural deviations in their trunk from the vertical. That response is called thigmomorphogenesis, and it is the ‘feedback correction’ to the more well-known gravitropism (the preference for growth vertically in the first place). Any bending stresses at the roots are amplified by the the fatigue induced by wind loading and thus must be minimised.
* Incidentally, John Perry, assistant to Lord Kelvin, was also the first to calculate a reasonably accurate estimate for the ancient age of the Earth, arriving at 2 billion years old - not bad for 1895!
An analysis by Greenhill in 1881 showed that an idealised pine tree of uniform trunk diameter 20 inches cannot grow beyond 90 m tall before failing by self-buckling. The actual tallest pine tree is about 83 m tall, which, for its 2 m diameter trunk base, is far below its theoretical limit. While the above Perry-Robertson model of imperfections might go some way to bringing the limit down, another limiting factor in tree height is the transpiration stream that nourishes the leaves, bringing the nutrients in the soil upwards through a constant flow of water. The suction pressure that drives this pump cannot be too strong, or the water high up in the xylem tubes would cavitate, stopping flow and starving the higher leaves of their craved electrolytes.
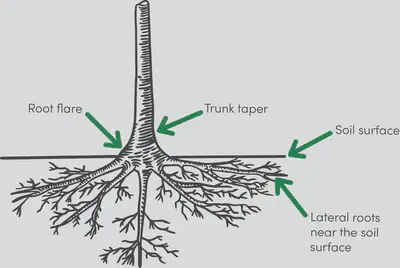
For most trees, it is in fact this hydrological constraint that limits maximum height, while structural stability guides development up to that limit. To eliminate the bending stresses induced in the base of the trunk due to swaying and leaning, trees have two interesting ways of strengthening themselves:
(1) vary the thickness of their trunk cross-section so that they are tapered (thicker at the base, thinner at the top), and
(2) reinforce their base on one side only by forming reaction wood to reduce peak stresses induced by the bending moment.
This motif of positive and negative feedback systems interacting to produce control-system-like functionality is a common one in biology, with perhaps the clearest examples being those of natural homeostasis (e.g. thermoregulation). Enrico Coen’s book Cells to Civilizations (2015) gives several further concrete examples, with a focus on how feedback system dynamics underpin development, the mechanistic basis for evo-devo biology. In (McMahon & Kronhauer, 1976) and (Kanahama & Sato, 2023), it is shown that the way trees taper their cross-sections (mechanism (1) above) is very close to the theoretically optimal distribution of trunk material. What looks like an intelligent design can therefore be explained by the optimising process of natural selection acting on heterotopy.
Where the common ancestry model shines
Let’s look further at mechanism (2) above, the production of reaction wood in response to asymmetric leaning. Reaction wood is produced by all woody plants, which includes trees. There are two types of reaction wood:
Compression wood, which is rich in the biopolymer lignin, is produced on the inner side of the leaning trunk (the side in compression), pushing up. Compression wood is the type of reaction wood produced in all gymnosperms (softwoods, non-flowering plants).
Tension wood, which is rich in the biopolymer cellulose, is produced on the outer side of the leaning trunk (the side in tension), pulling down. Tension wood is the type of reaction wood produced in most angiosperms (hardwoods, flowering plants).
.](/post/look-at-the-trees/reaction-wood_hue9b41c6be204dddf8328b45e28884cab_163256_60386ab7742b2eb29fb444ca411ea280.webp)
Seems like a clear-cut divide - the two big divisions of trees fit neatly into two different reaction wood types. Sounds like the beauty of design! But wait, what’s that sneaky word ‘most’ doing in that second point? As is all too common in biology, there’s always something that wants to be different…
Amborella is the only clade that bucks the trend: it’s an angiosperm that produces compression wood, like the gymnosperms, instead of tension wood, like its fellow angiosperms. At this point, we demand to know the answer to the following question:
“What is the evolutionary relationship between Amborella, the other angiosperms, and the gymnosperms?”
Evolution predicts nested hierarchies of traits. If the same trait pops up in multiple relatively unrelated groups of organisms, then although one mutation can be invoked to explain the trait in one such group, multiple independent mutations must be invoked to explain all of them. This is far less likely, and therefore has far weaker explanatory power, and is then shunted down the list of scientifically backed possibilities to explain the data at hand. This is where the principle of parsimony (more generally, Occam’s razor) is core to modern evolutionary theory. The phylogenetic tree structures recovered from genetic studies must at least somewhat match the trees deduced from more holistic observations, like traditional comparative anatomy. A counterexample to evolution’s parsimony is a way to falsify evolution (that’s your Precambrian bunny type of argument from Haldane).
So anyway, what’s the answer to the question above?
Molecular phylogenetic studies find that Amborella is the most evolutionarily basal extant angiosperm lineage (i.e. it is the sister clade to all other flowering plants). This divergence pattern is therefore entirely consistent with the predictions of evolutionary theory: the parsimonious conclusion is that the tension wood trait evolved once in the lineage leading to all other angiosperms after their split from Amborella. One new trait, one event in a single lineage - as tidy as it gets.
).](/post/look-at-the-trees/tree_evolution_hue651627ee9189f6c4fda8110ae805083_138410_66bbb22662b63b047857475ba9cc2815.webp)
Think about what it would take to explain this under a separate ancestry hypothesis. We are given that one type of tree is different to the others. Why? Since all separate ancestry models involve an omnipotent deity in the picture, such questions are frequently waved away with “mysterious ways”. But that’s not science. It’s not parsimonious. It has zero explanatory power. As Popper said,
“A theory that explains everything, explains nothing.”
If we try to steelman the modern intelligent design proponents’ argument, which is that because form and function are correlated, common functions reflect common design, not common descent, we still run into logical fallacies on their part. In biology, the causality is in the inverse direction of design: Crick’s central dogma is that function (what a protein does - function and phenotype) follows from form (protein shape, encoded by the DNA - genotype). The ID argument simply restates the observation as its own explanation (begging the question): it is a post hoc attribution of intentionalism that adds no new predictive content.
Branching out
There are also several poetic points about trees worth thinking about:
The ’tree’ of life
Evolution’s principal model is best illustrated by the tree of life, a highly cross-cultural symbol of origins. The data structure of the binary tree comes up in evolutionary theory because its recursive, ever-branching structure precisely mirrors the recursive, continuous process of speciation (ultimately the reason why cladistics replaced Linnaean taxonomy - there are no privileged ranks other than species, and even species can be ambiguous and is semi-arbitrary with its many species concepts).
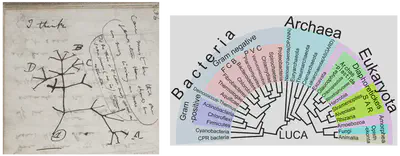
While there have been refinements to the idea of a single, perfectly bifurcating tree of life, such as the tree becoming more ‘bush-like’ near the last universal common ancestor (due to horizontal gene transfer among prokaryotes), or small mismatches at the tips of branches caused by incomplete lineage sorting (a transient consequence of genetic drift’s stochasticity), the underlying model remains powerful and broadly accepted.
Trees present a counter to irreducible complexity
Trees are climatic climax vegetation, the last and most mature stage of ecological succession, the process by which complex, interconnected, interdependent communities form in newly exposed land. This turns out to be a striking counterexample to the intelligent design (ID) proponents’ concept of irreducible complexity (IC): remove one species of an ecosystem and the food webs collapse, and yet we watch ecosystems form in real time. The interdependencies come later; they are not built in from the start. Complex traits in biology evolve in a similar way, with many direct examples known (e.g. the eye, the bacterial flagellum, the nervous system…), disproving IC, one of the core pillars of ID. ID was demonstrated and declared to be not scientific at the Kitzmiller v. Dover court case of 2005, with many of these examples of reducible complexity being key evidence.
.](/post/look-at-the-trees/ecological-succession_hu4e654550106ed520a938a4b579510f95_865740_dd037e5b0257f16372fd55bb2deca12f.webp)
Trees exemplify the thermodynamic purpose of life
Trees, like all photosynthetic life, are the primary producers for the vast majority of life*: they are responsible for capturing free energy and distributing it out to the rest of their biome’s food webs. Sunlight is the sole energy influx into the Earth, a natural free energy gradient that enables the development of non-equilibrium systems that will consume this free energy (high-exergy sunlight) and rapidly generate entropy irreversibly in the environment (water vapour from the transpiration stream output). This is the self-organising dissipative structuring principle that makes life compliant and specifically prompted by the laws of physics in the first place, as studied by many from Schrödinger (1944) to Schneider & Kay (1994) to Michaelian (2010s) to contemporary interdisciplinary discourse as reviewed in (Hall & McWhirter, 2023) and (Yong, 2025), with explanatory power in both abiogenesis and evolution.
.](/post/look-at-the-trees/thermo-life_hue0d47a340b90401e428b8daea35def1c_219685_8acac23677071ff4a1a1aa00c01f5cf2.webp)
* exception: chemosynthetic organisms living near deep hydrothermal vents, where sunlight cannot penetrate. Their free energy source is instead the geothermal heat of the Earth, provided via chemical-exergy-rich molecular fuel.
A ubiquitous property of life is that it is able to maintain its internal state, distinct to and despite disturbances from its environment (homeostasis). This is the precise opposite of thermodynamic equilibrium: any organism in equilibrium with its surroundings is dead. The energetic path to life is:
$$ \text{free energy input → nonequilibrium system} $$ $$ \text{→ dissipative structuring → simplest life} $$
Trees sit at both ends of this story. They are among the most complex macroscopic life forms on Earth, yet they remain directly coupled to the biosphere’s primary energy source, where nearly all other organisms have increasingly outsourced the collection of free energy to them. In doing so, they make the rest of the living world possible.
Conclusions
So, whether it’s the way trees grow structurally or energetically, we see optimisation that could be naively attributed to intelligent design. In a sense, it’s not wrong to say “look at the trees! they show design!” - BUT:
Design is incredibly hard to define rigorously, and counterexamples spring up as soon as we go beyond what’s intuitively known (which happens to be exactly the domain where we look to science for its powerful analytical toolkit, rather than relying on ‘common sense’ essentially invoked by ID)
Nature is more than capable of design! The constraints of biology, and the driving forces of chemistry and physics, work together to create ‘goal-oriented design’ (teleonomy) - selection for functionality, that is. We see the ‘goal’ because we’re intelligent, but we don’t invoke literal teleology because we also study the underlying causes.
The intuitive appeal of Paley’s watchmaker argument - that ID just puts a science-flavoured coat of paint on - funnels one into this fallacious line of reasoning, and is a logical chasm that Dennet separates clearly: from the initial ‘design stance’, one can either move to the ‘intentional stance’ or the ‘physical stance’. “Why do trees ‘want’ to grow tall? They must have been programmed to be tall!” No - it’s evolutionary dynamics. “Why do tree trunks support themselves so precisely? Obviously programmed.” No - it’s feedback system dynamics. “Why do trees ‘want’ sunlight at all? Programmed!” No - it’s thermodynamics! The existence of the designer isn’t disproven per se, their alleged actions are just made redundant in the natural world.
It could be argued that the role of the deity is not to create life directly, but to set up the laws of physics and initial conditions that make life inevitable. In my opinion, this Universe-scale fine tuning argument, valid or not, leads more towards theistic or deistic evolution rather than overt creationism or intelligent design - as evidenced by its support by most scientifically literate theists since the mid-20th century (e.g. Lemaître, founder of Big Bang cosmology) - and is therefore not contrary to the validity of evolution as a theory. The success of evolutionary algorithms in modern computer software demonstrates that when the rules of the game are intelligently designed, evolution follows naturally, with no further external intervention required as the world’s trajectory unfolds. If there is something behind it all, it must be unfathomably powerful; so powerful that one has to wonder whether it really cares as much about the trivialities of one species’ daily affairs on one planet in one galaxy living in one cosmic second as some assert it does.
Thanks for reading. As usual I intend to be mostly educational and mildly persuasive. Oh, and merry Christmas and a happy new year!
Lorcan Nicholls
Graduate Engineer
An graduate engineer from the University of Cambridge. Interested in interdisciplinary engineering and science, sustainable energy and automation.